דיאטה עתירת שומן מזרזת stemness וטומוריגניות של פרוגניטורים אינטסטינליים
קיים מידע מועט ביותר לגבי האופן שבו דיאטות הגורמות להשמנת יתר (אוביסיטי) מווסתות את התפקוד של תאי גזע רקמתיים ותאים פרוגניטוריים. מאמר זה מראה כיצד השמנת יתר הנגרמת מדיאטה עתירת שומן (HFD) מגדילה את המספר והתפקוד של תאי גזע אינטסטינליים Lgr5+ במעיים של יונקים. מבחינה מכאנית, HFD גורמת לחתימה איתנה של רצפטור דלתא המופעל על ידי פרוליפרטור פרוקסיזום (PPAR-δ) בתאי גזע אינטסטינליים ובתאים פרוגניטוריים (תאי גזע לא-אינטסטינליים), ואקטיבציה פרמקולוגית של PPAR-δ מובילה לרקפיטולציה של ההשפעות של HFD על תאים אלו. בדומה ל- HFD, טיפול ex vivo של תרביות אורגנואידים אינטסטינליים עם רכיבי חומצת השומן של HFD מזרז את פוטנציאל ההתחדשות העצמית של גופים אורגנואידים אלו באופן תלוי- PPARδ. באופן ראוי לציון, איתות PPAR-δ המופעל על ידי HFD ואגוניסטים מעניק יכולת אתחול אורגנואידים לפרוגניטורים, ואיתות PPAR-δ כפוי מאפשר לפרוגניטורים אלו ליצור גידולים in vivo לאחר אובדן מדכא הגידול Apc. ממצאים אלו מדגישים כיצד אקטיבציה של PPAR-δ באפנון-דיאטה משנה לא רק את התפקוד של תאי גזע אינטסטינליים ותאים פרוגניטוריים, אלא גם את היכולת שלהם לאתחל גידולים.
ידוע כי מעיים של יונקים מגיבים לאותות תזונתיים. תאי גזע אינטסטינליים (ISCs) Lgr5+ מעצבים מחדש את הרכב המעיים בתגובה לאותות בהשראת דיאטה, על ידי התאמת הייצור של תאי-בת ותאים פרוגניטוריים (תאים מגבירי-מעבר, שאינם ISCs), כאשר תאים פרוגניטוריים מתמיינים לסוגי התא השונים של המעיים. ה- Lgr5+ ISCs הגדולים נמצאים בבסיס הקריפטים של המעיים, בסמוך לתאי פאנט (paneth), המהווים רכיב עיקרי של נישת ה- ISC ומווסתים את ביולוגיית תאי הגזע בתגובה לדיאטה מוגבלת-קלוריות.
למרות שמחקרים אפידמיולוגיים ומחקרי מכרסמים חשובים מצביעים על קשר בין השמנת יתר להיארעות של סרטן המעי הגס, קיים מידע מועט בלבד לגבי האופן שבו האדפטציה של תאי גזע ותאים פרוגניטוריים לדיאטות מעודדות השמנת יתר משנה את הפוטנציאל של תאים אלו לאתחל גידולים. במעיים של עכברים, Lgr5+ ISCs מתפקדים בתור תאי המקור של לזיות אדנומטיות טרום-סרטניות הנגרמות בשל אובדן של הגן מדכא הגידול Apc; עם זאת, לא ברור האם מצב זה מתרחש בהקשר של טומרוגנזיס אינטסטנלי הקשור בהשמנת יתר. מאמר זה בודק כיצד השמנת יתר ארוכת טווח בהשראת HDF משפיעה על תפקוד תאי גזע אינטסטינליים ותאים פרוגניטוריים והמקור התאי של דיספלזיה אינטסטינלית.
HFD מגבירה את ספירת ה- ISC ואת תפקוד הקרפיטים
בכדי לאמוד את ההשפעות של השמנת יתר על ההומיאוסטאזיס של המעיים, שמרנו עכברים ב- HFD ארוכת טווח (דיאטה של 60% שומן, איור 1o extended data) במשך 9-14 חודשים, תקופה מספיקה בכדי להבחין ברבים מהפנוטיפים המטבוליים הקשורים בהשמנת יתר. באופן עקבי למחקרים קודמים, עכברים בתזונת HFD צברו מסה גדולה יותר באופן משמעותי בהשוואה לעכברים שקיבלו תזונה סטנדרטית (איור 1a extended data). בעוד שהמעי הדק של העכברים בתזונת HFD היה קצר יותר (איור 1c extended data) ושקל פחות (איור 1b extended data), לא נרשם שינוי בדחיסות של יחידות ה- crypt-villous (איור 1d extended data) או במספר של התאים האפופטוטיים (איור 1n extended data). מבחינה מורפולוגית, ה- HFD גרמה להפחתה קלה באורך ה- villi (איור 1g extended data), הפחתה קשורה במספר האנטרוציטים ב- villous (איור 1f extended data) והגדלה בעומק הקריפטים (איור 1e extended data). ה- HFD לא שינתה את המספר של תאי כרומוגרנין A+ אנטרואנדוקרינים או תאי גביע Alcian blue+ לכל יחידת crypt-villous של המעי הדק (איורים 2a-d extended data).
בכדי לבחון כיצד HFD משפיעה על התדירות של ISCs, ביצענו היברידיזציה in situ עבור olfactomedin 4 (olfm4), מארקר המתבטא על ידי Lgr5+ ISCs. בהשוואה לעכברים בתזונה רגילה, אלו שקיבלו תזונת HFD הפגינו עלייה של 50% במספר ה- olfm4+ ISCs (איור 1a ואיור 1I extended data). באופן מנוגד, ה- HFS הפחיתה את מספר תאי נישת הפאנט cryptdin 4+ (crp4+, ידועים גם כ- defa4+) ב- 23% (איור 1a ואיור 1m extended data). תצפיות אלו הובילו לשתי מסקנות: ראשית, HFD מזרזת את המספר ואת ההתחדשות העצמית של ה- ISC (לדוגמא, קריפטים עמוקים יותר עם יותר olfm4+ ISCs) על חשבון ההתמיינות (villi תאי קצר ומועט יותר); ושנית, ההתגברות של ISCs מתרחשת למרות ההפחתה במספר תאי הפאנט, ממצא המעלה את האפשרות כי תחת HFD, ה- ISCs מתאימים עצמם לפחות אינטראקציות מנישת תאי הפאנט שלהם.
בהתחשב בכך שהמספר והשגשוג של ISC גדלים ב- HFD (איור 1b, איורים 1h-k ו- 3f extended data ומידע נספח), בדקנו האם HFD מגבירה בנוסף את הרגנרציה של המעיים. באמצעות גישת in vitro, בחנו את היכולת של קריפטים אינטסטינליים מבודדים ליצור גופים אורגנואידיים בתרבית 3D. אורגנואידים אלו מובילים לרקפיטולציה של הארכיטקטורה האפיתליאלית והמגוון התאי של מעי יונקים ומשמשים כפרוקסי של פעילות ISCs, מכיוון שרק תאי גזע מסוגלים לאתחל ולשמר תאים אלו לטווח הארוך. קריפטים נגזרי-HFD מהמעי הדק והמעי הגס הפגינו סבירות גבוהה יותר לאתחל mini-intestines בתרבית, בהשוואה לאלו מהבקרה (איורים 1c, e, ואיור 3j extended data). בנוסף, אורגנואידים אלו היו ציסטיים יותר (כלמור, פחות ממוינים) במבנה שלהם והכילו פחות תחומי קריפטים (איור 1d). לאחר sub-cloning, אורגנואידים ראשוניים נגזרי-HFD חוללו יותר אורגנואידים משניים (איור 1f, איור 3k extended data). באופן עקבי לממצאים אלו, אורגנואידים נגזרי-קריפט של HFD הפגינו תדירות גבוהה יותר של Lgr5+ ISCs בהשוואה לבקרה (איורים 4a, d, e extended data), וכללו סוגי תא אינטסטינליים מגוונים, כמו תאי פאנט, ISCs, תאים אנטרואנדוקריניים ותאי גביע (איורים 4b-f extended data).
כדי לקבוע האם HFD מגבירה את הרגנרציה של הקריפטים in vivo, ביצענו הליך מיקרו-מושבות קלונוגני לזיהוי פעילות ISC. לאחר הקרנה בכמות קטלנית, עכברים בתזונת HFD הפגינו מספרים גבוהים של קריפטים שורדים ומשגשגים (Ki67+ תאים לכל קריפט) שכללו יותר olfm4+ ISCs לכל יחידת אורך של מעיים בהשוואה לבקרה (איורים 2e-g extended data). נתונים אלו תומכים בתפיסה לפיה HFD מעלה את המספר ואת יכולת הרגנרציה של ISCs in vitro ו- in vivo.
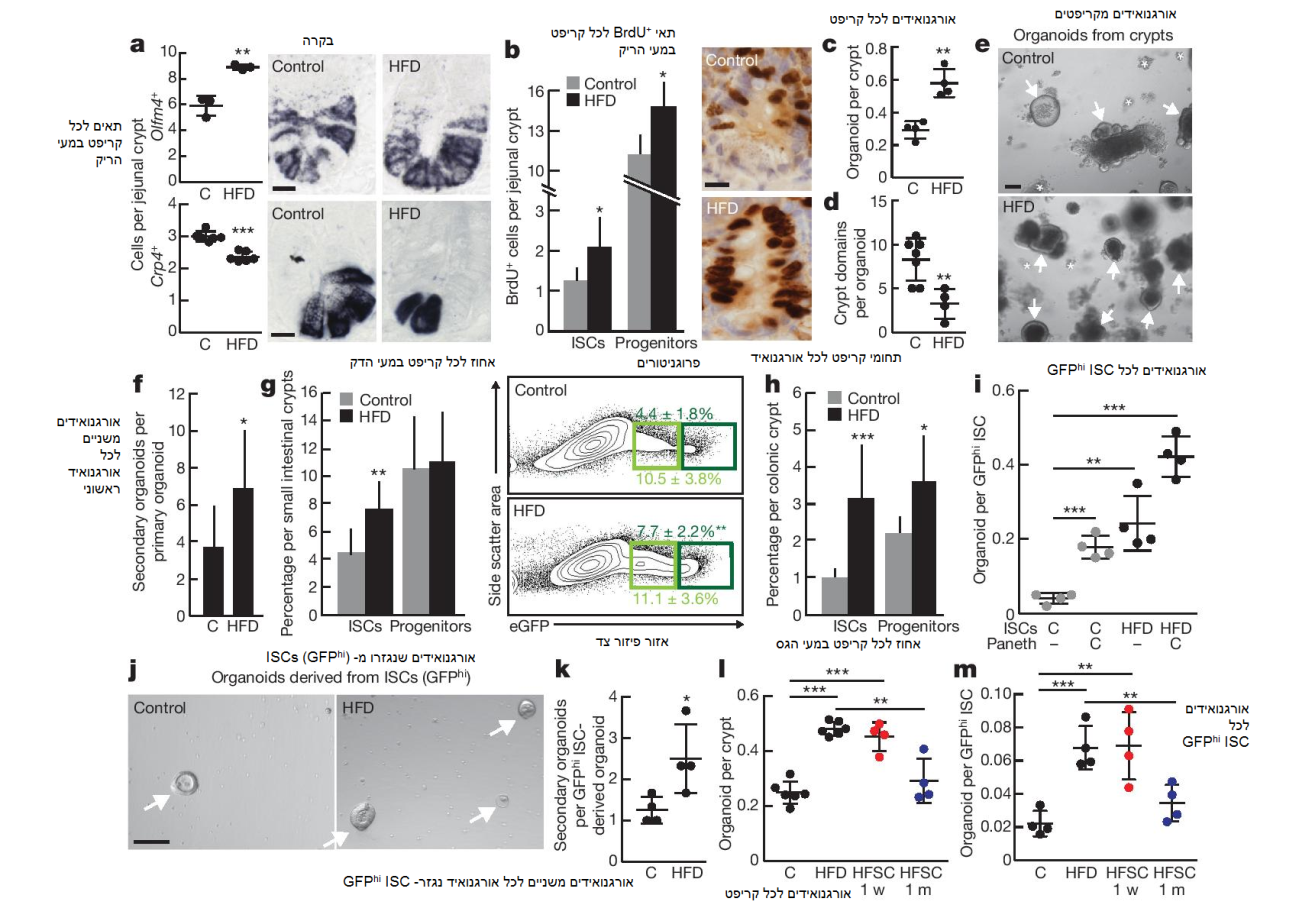
איור 1: HFD מגבירה את המספר והתפקוד של ISC. a) כימות של olfm4+ ISCs (n = 3) ותאי פאנט Crp4+ (n = 6) במעי הריק הפרוקסימלי של הבקרה (C) ועכברי HFD באמצעות היברידיזציה in situ. b) שילוב של BrdU ב- ISCs (תאים עמודיים של בסיס הקריפט) ובפרוגניטורים (תאים מגבירי-מעבר) לאחר פולס של 4 שעות (n = 6). c-e) כימות של אורגנואידים לכל קריפט (c, n = 4) ולכל תחום קריפט (d, n = 7) מהבקרה ומעכברי HFD. תמונה מייצגת: אורגנואידים יום 7 (e). החצים מסמנים אורגנואידים; כוכביות מסמנות קריפטים שהופסקו. f) מספר אורגנואידים משניים לכל אורגנואיד ראשוני נגזר-קריפט דיסוציאטיבי (n = 9 אורגנואידים ראשוניים, 3 אורגנואידים ראשוניים לכל דגימה עברו subcloning ב- 3 ניסויים נפרדים). g, h) תדירות של ISCs (Lgr5-GFPhi, ירוק כהה) ופרוגניטורים (Lgr5-GFPlow, ירוק בהיר) בכל המעי הדק (g, n = 10) ובמעי הגס (h, n = 8) לפי ציטומטריית זרימה. i, j) יכולת אתחול אורגנואידים של הבקרה ושל HFD ISCs בתירבות עם/ללא תאי פאנט (i, n = 4). תמונה מייצגת: אורגנואידים ראשוניים יום 5 (חצים, j). k) מספר אורגנואידים משניים לכל אורגנו איד משני דיסוציאטיבי נגזר-ISC (n = 4). l, m) קריפטים (l) ו- ISCs (m) שבודדו מעכברי HFD שהוחזרו לתזונה רגילה (HFSC) שימרו יכולת יצירת אורגנואידים גבוהה במשך שבוע אחד (w) (אדום; n = 4), אך לא במשך חודש אחד(m) (כחול; n = 4) בהשוואה לעכברי HFD מקבילים (n = 6 קריפטים, n = 4 ISCs). הנתונים הם ממוצע ± SD, אלא אם כן מצוין אחרת. * p < 0.05, ** p < 0.01, *** p < 0.001 (מבחן t סטודנטי). קנ”מ, 20 μm (a, b) ו- 100 μm (e, j). ניתוח היסטולוגי: a) olfm4: 10 קריפטים לכל קבוצה, crp4: 50 קריפטים לכל קבוצה; b) 50 קריפטים לכל קבוצה בכל ניסוי.
HFD מפחיתה את התלות בנישה של ISCs
על מנת לאמוד את ההשפעות של HFD על ISCs ועל הפרוגניטורים, השתמשנו בעכברי knock-in Lgr5-EGFP-IRES-CreERT2 עבור הכימות והבידוד של ISCs מבטאי חלבון פלורוסנטי ירוק (Lgr5-GFPhi) ותאים פרוגניטוריים (Lgr5-GFPlow). בהשוואה לבקרה, עכברי HFD הציגו תדירות מוגברת של Lgr5-GFPhi ISCs במעי הדק (איור 1g) ובמעי הגס (איור 1h, איור 3g extended data).
ההשפעות המנוגדות של HFD על מספרי ה- ISC ותאי הפאנט הובילו אותנו לשאול האם HFD משנה את תפקוד ה- ISC ואת התלות שלהם בנישה. בחנו את הפוטנציאל הקלונוגני של ISCs מעכברי הבקרה ומעכברי ה- HFD לבדם, או יחד עם תאי נישת הפאנט. באופן עקבי למחקרים קודמים, ה- ISCs של הבקרה, בפני עצמם, לא יצרו אורגנואידים בכמות מספקת, אך יצרו אורגנואידים במידה איתנה בתירבות משותף עם תאי פאנט (איור 1i). באופן מפתיע, ISCs נגזרי-HFD לבדם (ללא תאי פאנט) הציגו יכולת מוגברת לאתחל אורגנואידים עם התמיינות מרובת-שורות (multilineage) ויותר אורגנואידים משניים בהשוואה ל- ISCs של הבקרה (איורים 1i-k, איורים 4h, i, l, m extended data). התירבות המשותף עם תאי פאנט הגביר את פעילות אתחול האורגנואידים של HFD ISCs (איור 1i). אורגנואידים שנגזרו מהבקרה ומ- HFD ISCs לבדם ייצרו באופן אפקטיבי תאי פאנט בתוך 24 שעות מתחילת התירבות (איורים 4j, k extended data). בנוסף, קריפטים ו- ISCs שבודדו מעכברים שניזונו מ- HFD אך הוחזרו לדיאטה רגילה, שימרו יכולת מוגברת לאתחול אורגנואידים במשך יותר מ- 7 ימים אך פחות מ- 4 שבועות, ממצא המלמד כי ההשפעות של HFD הן הפיכות (איור 1l, m). נתונים אלו, לצד הממצא כי HFD גורם ל- uncoupling של התרחבות in vivo של ISCs מנישת תאי הפאנט שלהם, מלמדים כי ISCs עוברים שינויים אוטונומיים בתגובה ל- HFD הממצבים אותם לגדילה שאינה תלוית-נישה בהליך (assay) האורגנואידים.
חומצות שומן מניעות התחדשות עצמית של האורגנואידים
על מנת לבדוק האם הרכיבים הדיאטטיים של HFD יכולים לגרום לרקפיטולציה של אספקטים של פנוטיפ תאי הגזע שנגרם מ- HFD, הרחבנו את אורגנואידי הבקרה בתווך קריפטים בתוספת חומצה פלמיטית, רכיב עיקרי של HFD. הטיפול בחומצה הפלמיטית לא שינה את הפוטנציאל הקלונוגני של קריפטי הבקרה בתרבית הראשונית (איור 2a). עם זאת, בדומה לאורגנואידים מעכברי תזונת ה- HFD, האורגנואידים הראשוניים שנחשפו ex vivo לחומצה פלמיטית הציגו יותר אורגנואידים משניים בהשוואה לבקרה (איורים 2b, c, איור 5a extended data). באופן עקבי לממצאים אלו, האורגנואידים שטופלו עם חומצה פלמיטית כללו כמעט פי 2 יותר Lgr5+ ISCs (איור 2d, איור 5b extended data), והפגינו תלות מופחתת בנישה בהליך האורגנואידים (איור 2e). תוצאות דומות הושגו עם חומצות שומן אחרות כמו חומצה אולאית ותערובת ליפיד באורגנואידים אינטסטינליים של עכברים ובני אדם (איור 3h-k, איור 5c-f extended data). ממצאים אלו מלמדים כי רכיבי דיאטה עיקריים של HFD הינם מספקים בכדי ליצור רקפיטולציה של אספקטים של פנוטיפ תאי הגזע של HFD in vivo.
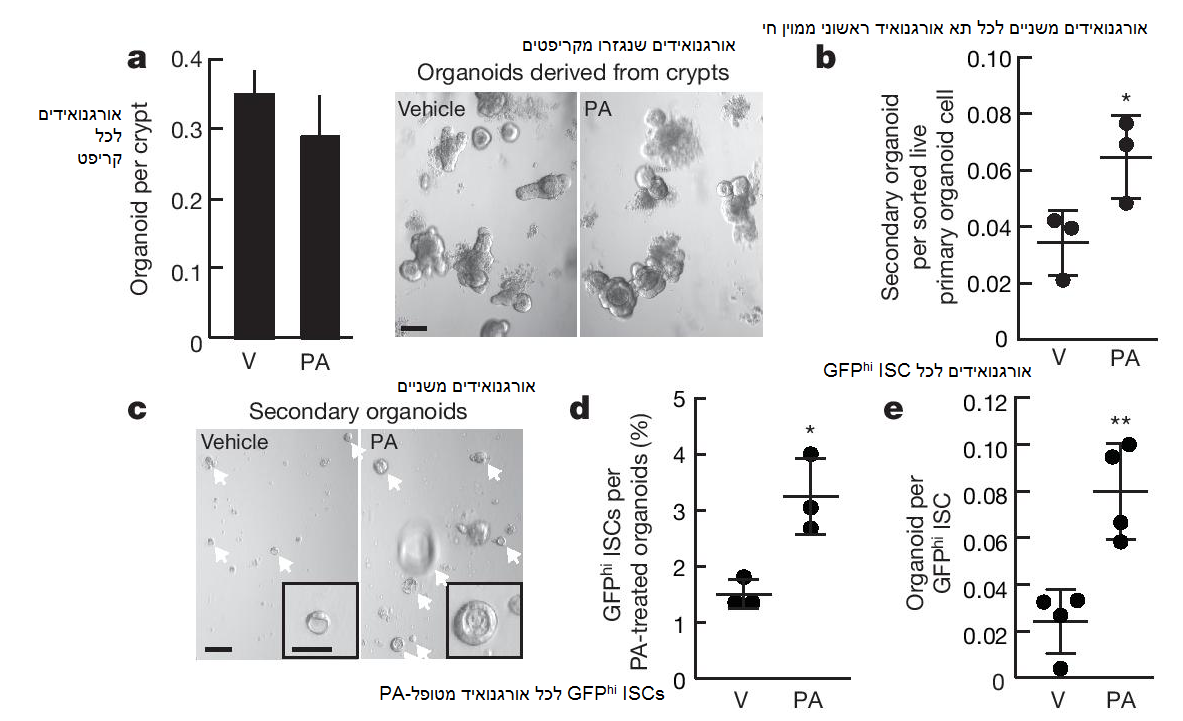
איור 2: חשיפה ex vivo של אורגנואידים אינטסטינליים לחומצה פלמיטית יוצרת רקפיטולציה של אספקטים של HFD. a) קולונוגניות של קריפטים נאיביים שתורבתו עם 30 μM חומצה פלמיטית (PA) בתרביות אורגנואידים ראשוניים (n = 5). תמונה מייצגת: אורגנואידים יום 4. V, vehicle. b, c) יצירת אורגנואידים משניים של 1000 תאי אורגנואידים ראשוניים ממוינים חיים לאחר 4 שבועות של טיפול 30 μM חומצה פלמיטית (b, n = 3). תמונה מייצגת: אורגנואידים משניים יום 4 (חצים, c). d, e) תדירות (d) ואתחול אורגנואידים (e) של ISCs (Lgr5-GFPhi) לאחר 4 שבועות של חשיפה ל- 30 μM חומצה פלמיטית (d, n = 3; e, n = 4). הנתונים הם ממוצע ± SD אלא אם מצוין אחרת, מ- n ניסויים נפרדים. *p < 0.05, ** p < 0.01 (מבחן t סטודנטי). קנ”מ, 100 μm (a, c) ו- 50 μm (c, inset).
HFD פועלת דרך PPAR-δ ב- ISCs
בכדי לקבל תובנות מכאניות לגבי האופן שבו HFD מתווכת השפעות אלו, ביצענו ריצוף messenger RNA על Lgr5-GFPhi ISCs ותאי פרוגניטור Lgr5-GFPlow מעכברי הבקרה ועכברי ה- HFD, בהתאמה (איור 6p extended data). נתיב ניתוח העשרת סט גנים (GSEA) וניתוחי מוטיב קשירת גורמי שעתוק חשפו העשרה של מטרות השעתוק ומוטיבי הקשירה של משפחת רצפטור הגרעין המופעל על ידי פרוקסיזום פרוליפרטור (PPAR) ו- X רצפטור כבד/רטינואיד של שותפי הקשירה ההטרודימריים של PPAR (LXR/RXR; איורים 6c, d extended data). שלושה חברים מרכיבים את משפחת ה- PPAR (α, δ ו- γ); מביניהם, PPAR-δ (Ppard) הוא הפרה-דומיננטי המתבטא בתאי גזע אינטסטינליים ותאי פרוגניטור ברמת ה- mRNA בעכברי ה- HFD והבקרה (איורים 6a, b extended data). לפיכך, בחרנו להתמקד ב- PPAR-δ והתפקיד הפוטנציאלי שלו בצימוד של HFD ל- ISC.
למרות שביטוי ה- PPAR-δ כשלעצמו לא גבר באופן משמעותי (איורים 6a, b extended data), ה- HFD גרמה לביטוי איתן של רבים מגני המטרה שלה ברמות ה- mRNA הן במעי הדק (איור 6e extended data) והן במעי הגס (איור 3h extended data). ההשראה של תוכנית ה- PPAR-δ אומתה ברמת החלבון ב- ISCs ובפרוגניטורים (איור 3a). על מנת לבדוק פונקציונאלית האם תוכנית PPAR-δ יכולה לחקות את HFD, יישמנו את אגוניסט ה- PPAR-δ GW501516 במשך 4 שבועות על עכברי Lgr5-EGFP-IRES-CreERT2. הטיפול הוביל להשראה חזקה של חלבוני המטרה של PPAR-δ ב- ISCs ובפרוגניטורים (איור 3a). בנוסף, איתות PPAR-δ מופעל-אגוניסט הגביר את תדירויות in vivo של olfm4+ ו- Lgr5+ ISCs (איורים 3b, e, איור 6f extended data) ואת השגשוג של תאי גזע ותאי פרוגניטור (איור 3c), אך ללא השפעה על מספר תאי פאנט (איור 3b, איור6g extended data). ראוי לציין כי קריפטים במעי הדק (איור 3d) ובמעי הגס (איור 3l, m extended data) מעכברים שטופלו באגוניסט אתחלו יותר אורגנואידים בהשוואה לאלו שטופלו ב- vehicle. בדומה ל- ISCs מעכברי HFD, ה- ISCs שנגזרו מעכברים שטופלו באגוניסט היו אפקטיביים יותר באתחול אורגנואידים ללא תלות בתאי פאנט, בהשוואה לבקרה (איור 3f). בנוסף, אורגנואידים שנחשפו לאגוניסט ה- PPAR-δ כללו יותר Lgr5+ ISCs (איור 3g) ויכולת התחדשות עצמית גבוהה יותר בהליכים משניים (איור 3i). נתונים אלו מלמדים כי איתות PPAR-δ ממושך יוצר רקפיטולציה של השפעות ה- HFD על תפקוד ה- ISC.
מכיוון שחומצות שומן ex vivo מחקות אספקטים של HFD, בדקנו האם תופעה זו מתרחשת דרך איתות PPAR-δ. בדומה ל- HFD, מצאנו כי חשיפה ex vivo של אורגנואידים של עכברים ובני אדם לחומצות שומן מעוררת תכנית PPAR-δ איתנה (איורים 5g-j ו- 6h extended data). על מנת לאמוד את הנחיצות של PPAR-δ בתגובה זו לחומצות השומן, יצרנו עכברי Ppard ספציפי למעיים, בהשראת טמוקסיפין (איורים 6i, j extended data). אבלציה אקוטית של Ppard במעיים הייתה ללא השפעה ניכרת על המספר, השגשוג או התפקוד של ISCs ותאי פרוגניטור (איור 3h, איור 6i-n extended data). עם זאת, אובדן ה- Ppard עצר גם את ההשפעות מזרזות ההתחדשות העצמית של חומצות השומן ואגוניסט ה- PPAR-δ, וגם את ההשראה של ביטוי גן המטרה PPAR-δ בהליכי האורגנואידים המשניים (איור 6o extended data). ממצאים אלו מראים כי PPAR-δ מתווך את ההתחדשות העצמית של אורגנואידים בהנעת חומצות שומן.
HFD ו- PPAR-δ מגבירים פעילות של β-catenin
מכיוון שאקטיבציה שלHFD ו- PPAR-δ מעבירה תפקוד מוגבר של תאי גזע, בחנו האם התערבויות אלו מווסתות את נתיב Wnt/β-catenin, הדרוש לתחזוקת ISC. ראשית, מצאנו יותר β-catenin גרעיני, פרוקסי לפעילותו, ב- ISCs ופרוגניטורים ממוינים ועל סקציות אינטסטינליות מעכברי HFD ועכברים מטופלי אגוניסט PPAR-δ בהשוואה לבקרה (איורים 7c-i extended data). שנית, קריפטים מעכברי HFD ועכברים מטופלי אגוניסט PPAR-δ דרשו פחות Wnt אקסוגני עבור תחזוקת האורגנואידים בהשוואה לבקרה (איורים 4a, b). לבסוף, מצאנו כי רמות מוגברות של β-catenin גרעיני קשורות ל- PPAR-δ בקריפטים של HFD (איורים 7j-l extended data).
בכדי לבדוק כיצד HFD ו- PPARδ מופעל-אגוניסט משפיעים על פעילות השעתוק של β-catenin, ביצענו מולטיפלקס PCR שעתוק לאחור כמותי חד-תאי על בסיס מיקרופלואידי (qRT-PCR) באמצעות פריימרים עבור רשימה נבחרת של גני מטרה β-catenin ידועים הכוללים מארקרים של ISCs (טבלה 2 נספחים, איורים 8a, d extended data). בעוד ש- HFD לא שינתה את הביטוי של גני החתימה של תאי גזע (כלומר, Lgr5) המשתנים בין תאי גזע לבין תאי פרוגניטור (איורים 8b, e, g, h extended data), היא עוררה ביטוי של תת-קבוצה של גני מטרה β-catenin כמו Bmp4, Jag1, Jag2 ו- Edn3 ב- ISCs ובפרוגניטורים (איורים 4c, e, איורים 3i ו- 8c, f, I, j extended data, מידע נספח). ניתוח qRT-PCR חד-תאי אישר כי PPAR-δ מופעל-אגוניסט גרם בנוסף להשראת שעתוק של Bmp4, Jag1, Jag2 ו- Edn3 ב- ISCs ובפרוגניטורים (איורים 4d, f). אימתנו בנוסף את ביטוי Jag1 באמצעות היברידיזציית מולקולה אחת in situ ומצאנו כי הוא מתבטא באופן נרחב בתוך HFD (איור 8k extended data). בנוסף, בתגובה לחומצות שומן in vitro, ה- PPAR-δ היה נחוץ עבור ההשראה של Jag1 ו- Jag2 באורגנואידים משניים (איור 6o extended data). בסך הכול, תוצאות אלו תומכות במודל לפיו HFD מפעילה תת-קבוצה של גני מטרה β-catenin בתיווך PPAR-δ ב- ISCs ותאי פרוגניטור.
כדי לבדוק האם קיימת תכנית דומה במודל חלופי של השמנת יתר, אמדנו כיצד המעיים מסתגלים להשמנת יתר אצל עכברים בעלי חוסר ברצפטורי לפטין (db/db) – מודל השמנת יתר המתפתח על דיאטה רגילה. בסך הכול, מצאנו כי האדפטציה האינטסטינלית במודל ה- db/db של השמנת היתר היה ברובו הפוך לממצאים של עכברי ה- HFD (איור 9 extended data, מידע נספח). הבדלים כאלו מראים כי אפילו במקרה של השמנת יתר, הדיאטה משפיעה על הביולוגיה של ISC ופרוגניטורים.
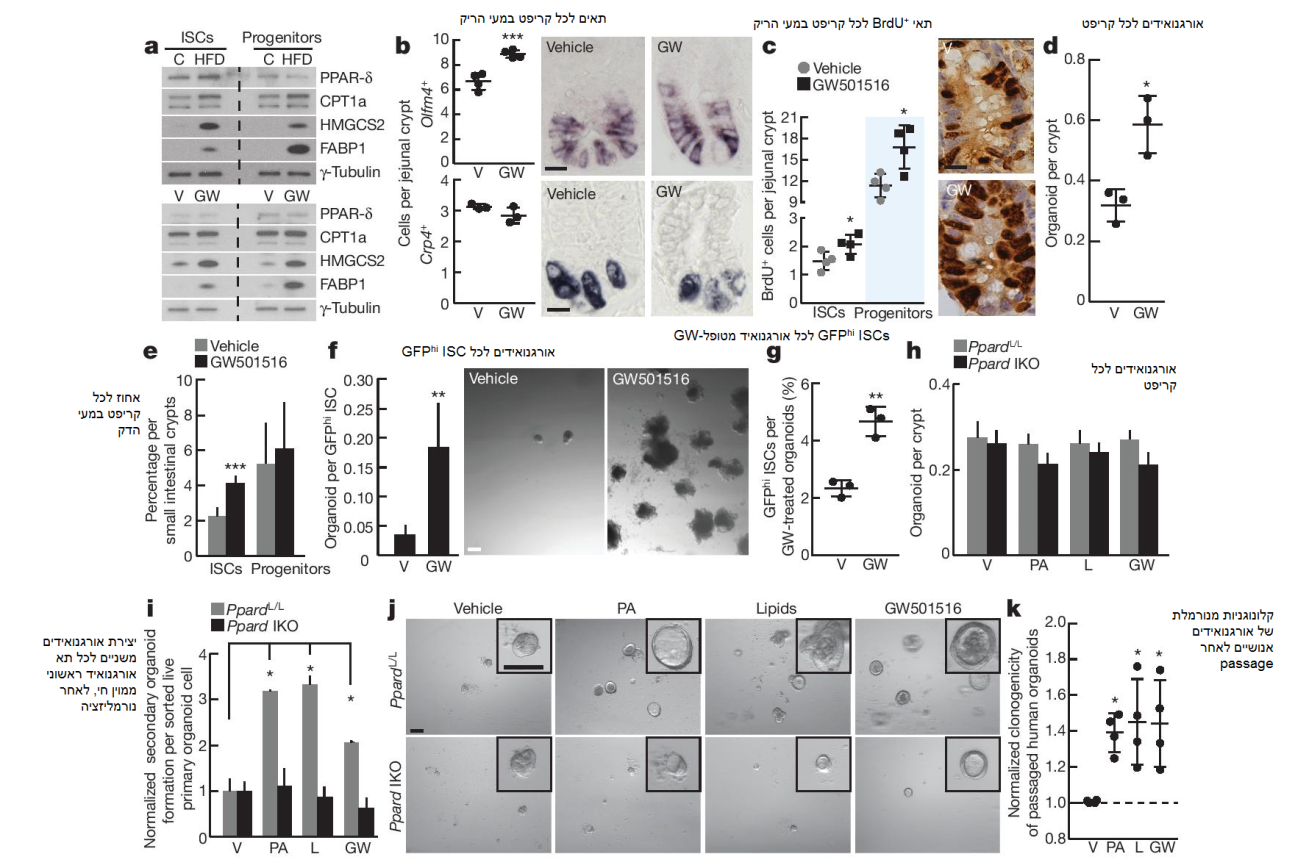
איור 3: PPAR-δ אקטיבי ב- ISCs מתווך את ההשפעות של HFD. a) אימונובלוטים של חלבוני מטרה PPAR-δ ב- ISCs (Lgr5-GFPhi) ופרוגניטורים (Lgr5-GFPlow) ממוני-זרימה מעכברי בקרה, HFD, vehicle ו- GW501516 (GW) (n = 2). b) כימות של olfm4+ ISCs (n = 4) ותאי פאנט Crp4+ (n = 3) באמצעות היברידיזציה in situ בקריפטים פרוקסימליים במעי הריק. c) שילוב BrdU ב- ISCs (תאים עמודיים של בסיס הקריפט הסמוכים לתאי פאנט) ובפרוגניטורים (תאים מגבירי-מעבר שאינם סמוכים לתאי פאנט) לאחר פולס של 4 שעות (n = 4). d) אורגנואידים לכל קריפט מעכברי vehicle ועכברי טיפול- GW501516 (n = 3). תדירויות של ISCs (Lgr5-GFPhi) ופרוגניטורים (Lgr5-GFPlow) ממויני-זרימה (n = 5) מכל המעי הדק של עכברי vehicle ועכברי טיפול- GW501516. f) יכולת אתחול אורגנואידים של ISCs שנגזרו מעכברי vehicle ומעכברי טיפול- GW501516. תמונה מייצגת: אורגנואידים יום 12 (n = 5). g) תדירות של ISCs (Lgr5-GFPhi) באורגנואידים לאחר 14 יום של חשיפה ל- GW501516 ex vivo (n = 3). h-j) יכולת יצירת אורגנואידים ראשוניים (h, n = 5) ומשניים (I, j, n = 5; מנורמל ל- vehicle) של עכברי בקרה ( PpardL/L (L, loxP)) ועכברי Ppard knockout אינטסטינלי (IKO) בעת טיפול ex vivo עם תערובת vehicle, חומצה פלמיטית וליפיד (L) ו- GW501516. תמונה מייצגת: אורגנואידים משניים יום 4 (j). k) קלונוגניות מנורמלת של אורגנואידים אינטסטנליים אנושיים לאחר טיפול ex vivo עם תערובת חומצה פלמיטית וליפיד ו- GW501516 בתרבית משנית (n = 4, ראה שיטות). הנתונים הם ממוצע ± SD אלא אם מצוין אחרת, מ- n ניסויים נפרדים. * p < 0.05, ** p < 0.01, *** p < 0.001 (מבחן t סטודנטי). קנ”מ, 20 μm (b, c), 200 μm (f) ו- 100 μm (j). ניתוח היסטולוגי: b, olfm4: 15 קריפטים לכל קבוצה, crp4: 50 קריפטים לכל קבוצה; c, 50 קריפטים לכל קבוצה בכל ניסוי.
PPAR-δ מאפשר גידולים אצל non-ISCs
תאי גזע סומאטיים לעתים קרובות צוברים את המוטציות הראשוניות המובילות לשינויים אונקוגניים. מצאנו כי ב- HFD ישנה שכיחות גדולה יותר במעיים של לזיות דיספלסטיות ספונטניות ברמה נמוכה (אדנומות), קרצינומות, או שתיהן, בהשוואה לבקרה (איורים 5a-c), נתון שעשוי לשקף את העובדה כי קיימים יותר ISCs בעכברי HFD המסוגלים לרכוש מוטציות אונקוגניות. מכיוון ש- PPAR-δ בהשראת-HFD גורם בנוסף לאקטיבציה של חתימת β-catenin בתאים פרוגניטוריים, הצענו כי אוכלוסיות פרוגניטורים non-ISCs יכולות לרכוש תכונות של תאי גזע ולתרום לאתחול גידולים במקרים של השמנת יתר הנגרמת מדיאטה. כדי לבחון אפשרות זו, בדקנו האם HFD או PPAR-δ מופעל-אגוניסט השפיעו על יכולת אתחול האורגנואידים של הפרוגניטורים (non-ISC). מעניין לציין כי פרוגניטורים Lgr5-GFPlow, אך לא אנטרוציטי villous ממוינים טרמינאלית, מעכברי HFD ועכברי PPAR-δ מטופל-אגוניסט יצרו אורגנואידים (איורים 5d, e, ואיורים 4g, 7a, b extended data), ממצא המצביע על האפשרות כי איתות PPAR-δ כפוי בפרוגניטורים אינטסטינליים לא רק מעניק יכולת אתחול אורגנואידים אלא גם פוטנציאל אתחול גידולים.
על מנת לבחון אפשרות זו, יצרנו עכברי ApcL/L; Lgr5-EGFP-IRES-CreERT2 כדי לאמוד האם אקטיבציה פרמקולוגית של PPAR-δ מאפננת את היכולת הטומוריגנית של פרוגניטורי Lgr5-GFPlow. זריקת טמוקסיפן מובילה לאובדן Apc ב- Lgr5-GFPhi ISCs, אשר בתורם מחוללים פרוגניטורים Lgr5-GFPlow שהם Apc-null (איור 10c extended data). 4 ימים לאחר מתן הטמוקסיפן, בודדנו את ה- Apc-null Lgr5-GFPhi ISCs ואת הפרוגניטורים Lgr5-GFPlow מה- vehicle ומהעכברים מטופלי PPAR-δ-אגוניסט כדי לאמוד את פוטנציאל יצירת הגידולים של אוכלוסיות אלו באמצעות הליכים נפרדים (איור 5f, סכמה): ראשית, בחנו את היכולת שלהם לגרום ליצירת אורגנואידים אדנומטיים בתרבית; שנית, בחנו את היכולת של 10,000 תאי גזע ותאי פרוגניטור ממוינים לאתחל אדנומות במעי גס סינגנאים.
מצאנו כי Apc-null ISCs מעכברים מטופלי PPAR-δ-אגוניסט היו קלונוגניים במידה זהה לאלו של הבקרה; עם זאת, איתות PPAR-δ כפוי בפרוגניטורים Apc-null הגביר את יכולתם ליצור אורגנואידים אדנומטיים באופן ניכר (איור 5g). בשלב הבא, בדקנו את הפוטנציאל של ISC ותאי פרוגניטור שבודדו לאחרונה ליצור אדנומות אינטסטינליות in vivo (איור 5f, סכמה). בדומה להליך האורגנואידים, לאגוניסט ה- PPAR-δ לא הייתה השפעה מוספת על היכולת של תאי גזע שהם Apc-null ליצור אדנומות β-catenin+ (Apc-null) (איור 5h, איורים 10a, d estended data) בהשוואה לבקרת ה- vehicle. עם זאת, איתות ה- PPAR-δ הכפוי אפשר לפרוגניטורים Apc-null, אך לא למקבילים מטופלי ה- vehicle שלהם, לאתחל אדנומות β-catenin+ (Apc-null) באופן איתן לאחר ההשתלה במעי הגס (איור 5h, איורים 10b, d extended data). נתונים אלו מלמדים כי אקטיבציה של PPAR-δ מאפשרת לתת-קבוצה של פרוגניטורים non-ISCs לאתחל גדילה אדנומטית in vitro ו- in vivo (ראה מידע נספח).
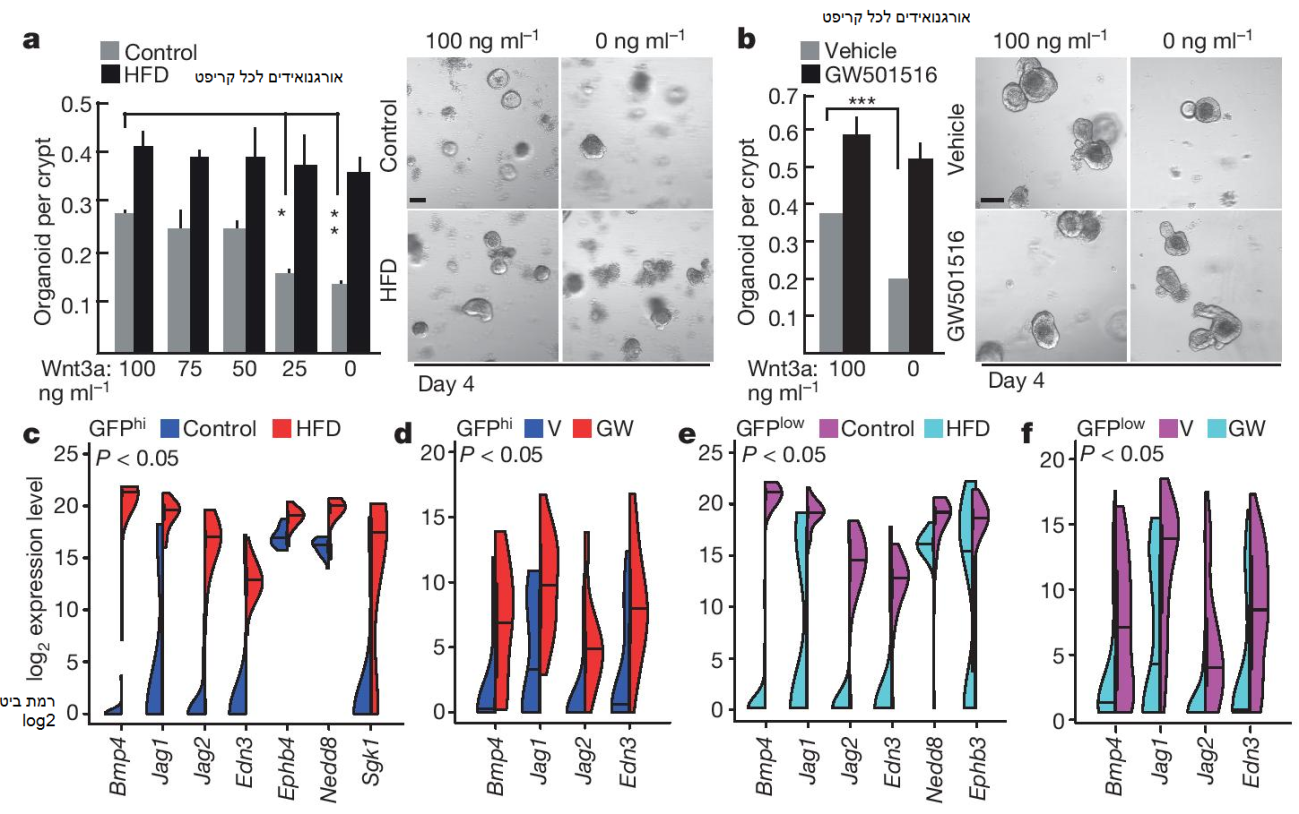
איור 4: איתות PPAR-δ בהשראת-HFD מביא לביטוי של תת-קבוצה של גני מטרה β-catenin. a, b) כימות אורגנואידים לכל קריפט מעכברי HFD (a) ועכברי GW501516 (b) עם ריכוזים של Wnt3a. תמונה מייצגת: אורגנואידים 4 יום (n = 5). c-f) מתווה violin עבור גני המטרה β-catenin עם ההשראה הגבוהה ביותר ב- ISCs (Lgr5-GFPhi) מעכברי HFD (c) ועכברי טיפול-GW501516 (d) (24 תאים בודדים לכל קבוצה), ובפרוגניטורים (Lgr5-GFPlow) מעכברי HFD (e) ועכברי טיפול-GW501516 (f) (72 תאים בודדים לכל קבוצה). הנתונים הם ממוצע ± SD אלא אם מצוין אחרת, מ- n ניסויים נפרדים. * p < 0.05, ** p < 0.01, *** p < 0.001 (מבחן t סטודנטי). קנ”מ, 100 μm.
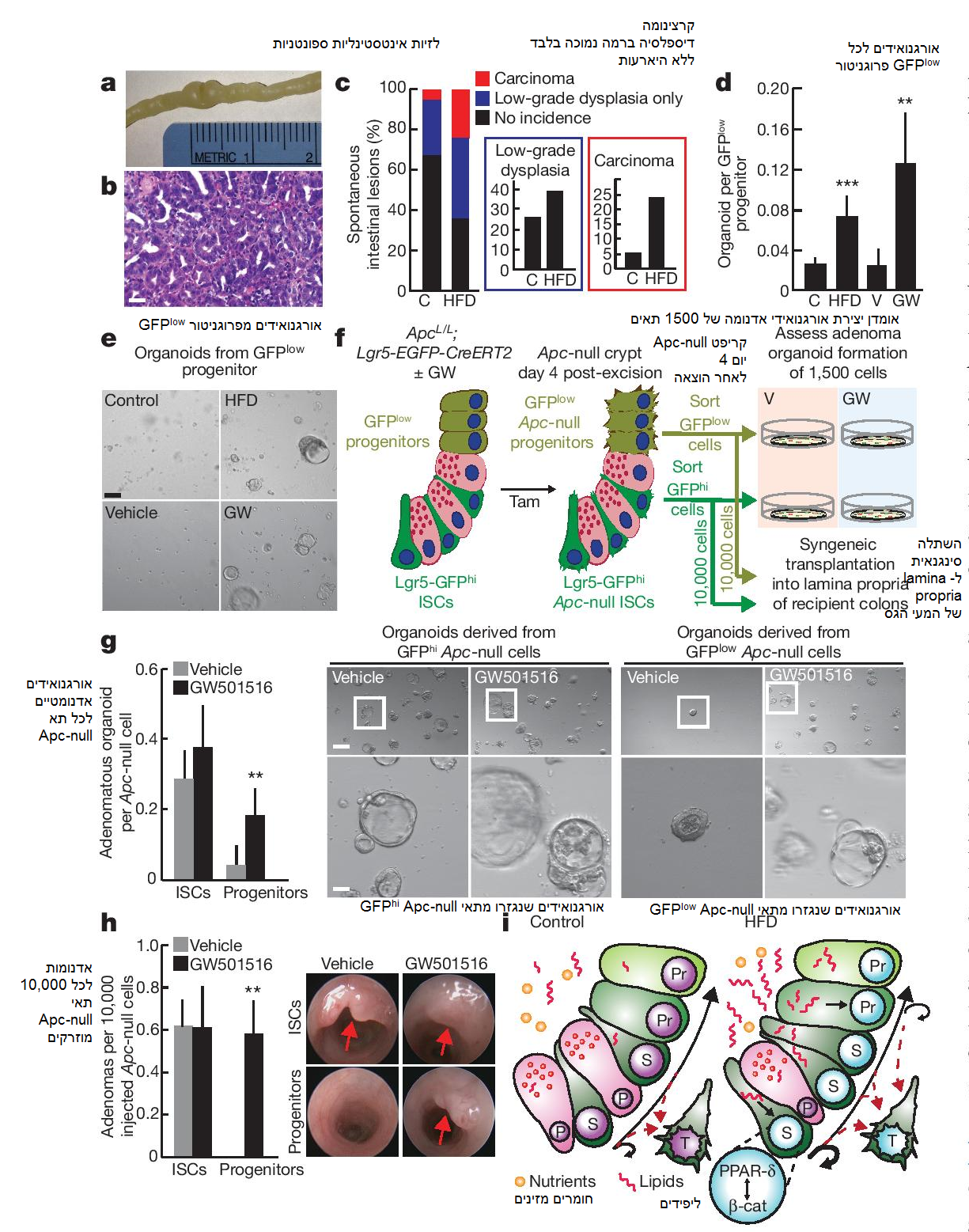
איור 5: אקטיבציה של PPAR-δ מעבירה יכולת אתחול אורגנואידים וגידולים לתאים שאינם-גזע. a, b) גידול אינטסטינלי ספונטני מייצג מעכבר HFD: תמונה כללית (a) ותמונת המטוקסילין ואאוזין (H&E) מיקרוסקופית (b). c) היארעות של לזיות ספונטניות אינטסטינליות דיספלסטיות ברמה נמוכה (אדנומה) וקרצינומות בעכברי הבקרה (n = 19) ועכברי HFD (n = 25). d, e) יכולת אתחול אורגנואידים של פרוגניטורים (Lgr5-GFPlow) מעכברי HFD (n = 7) ועכברי GW501516 (n = 5). תמונה מייצגת: אורגנואידים יום 7 (e). f) אומדן סכמטי in vitro ו- in vivo של יכולת אתחול אדנומה של Apc-null ISCs (Lgr5-GFPhi) ופרוגניטורים (Lgr5-GFPlow) מעכברי vehicle ועכברי GW501516. Tam, טמוקסיפן. g) מספרים ותמונות יום 5 מייצגות של אורגנואידים אדנומטיים מ- Apc-null ISCs (Lgr5-GFPhi) ופרוגניטורים (Lgr5-GFPlow) שטופלו עם/ללא GW501516 בתווך EN (EGF ו- noggin בלבד) (n = 6). h) קולונוסקופיה אופטית של גידולים שנוצרו לאחר השתלה אורטוטופית של 10,000 Apc-null ISCs (Lgr5-GFPhi) או פרוגניטורים Apc-null (Lgr5-GFPlow) מעכברי vehicle ועכברי GW501516 (n = 5). i) מודל של אדפטציה אינטסטינלית ל- HFD: מכאנית, ה- HFD מפעילה תוכנית בתיווך-PPAR-δ המגבירה את יכולת אתחול האורגנואידים והגידולים של פרוגניטורים אינטסטינליים. אחת התכונות של תוכנית ה- PPAR-δ כוללת השראה של תת-קבוצה של גני מטרה β-catenin. P, תא פאנט; Pr, תא פרוגניטור; S, תא גזע; T, תא גידולי. קווי נקודות אדומות מסמנים תאי Apc-null עם יכולת יצירת גידולים. הנתונים הם ממוצע ± SD אלא אם מצוין אחרת, מ- n ניסויים נפרדים. * p < 0.05, ** p < 0.01, *** p < 0.001 (מבחן t סטודנטי). קנ”מ, 50 μm (b, g למטה) ו- 200 μm (e, g למעלה).
דיון
הנתונים שלנו מצביעים על מודל לפיו HFD מגבירה התחדשות עצמית של ISC ונותנת תכונות של stemness (כלומר, יכולת אתחול אורגנואידים) על פרוגניטורים שאינם-תאי גזע על ידי אקטיבציה של איתות PPAR-δ (איור 5i). מחקר קודם מראה כי משטר דיאטה שונה, הגבלת קלוריות, מגביר את מספר תאי הגזע ותאי פאנט ומווסת את תפקוד ה- ISC הלא-תאי אוטונומית דרך נישת תא פאנט, ללא השפעה מובהקת על תפקוד תאי הפרוגניטור. באופן מנוגד, במחקר זה מצאנו כי HFD ארוכת טווח מביאה להשפעות מנוגדות על מספר תאי הגזע ותאי פאנט, וכי תאי גזע אלו תלויים פחות בתאי הפאנט בהליכים תפקודיים. העובדה כי מצאנו השראה של מטרות ה- β-catenin Jag1 ו- Jag2 (אופיינית, ה- notch ligands מתרחבים באמצעות תאי פאנט) בתאי גזע ותאי פרוגניטור ב- HFD מצביעה על תפקיד אפשרי של איתות notch. ב- HFD, ISCs או תאי פרוגניטור סמוכים עשויים לתפקד בתור מקור חלופי של notch ligands עבור Lgr5+ ISCs שאינם במגע ישיר עם תאי פאנט, ומאפשרים להם להתמיד in vivo ובהליך האורגנואידים. מחקר עדכני המראה כי אקטיבציה של PPAR-δ בעצמות מגבירה את איתות ה- β-catenin הינו עקבי לממצא שלנו כי PPAR-δ מופעל-דיאטה מפעיל תכנית β-catenin מוגבלת. גנים בתוך תכנית ה- β-catenin מופעלת-PPAR-δ כוללים Jag1, Jag2 ו- Bmp4, שלעתים קרובות עוברים דה-רגולציה בטומוריגנזיס אינטסטינלי מוקדם (איורים 4c-f).
המחקרים האחרונים מציעים כי גורמים אינטרינזיים ואקסטרינזיים תורמים לסיכון לסרטן באמצעות שליטה של חלוקות תאי גזע. מודלים אלו מנבאים כי גורמים אינטרינזיים כמו HFD עלולים להגדיל את הסיכון לסרטן על ידי הגברת חלוקות תאי-גזע, שהם תאי המקור של סוגי סרטן רבים. הנתונים שלנו (איור 5i) ומחקרים קודמים מראים כי HFD מגבירה את המספר והשגשוג של ISCs, שעשויים להיות אחראים חלקית לעלייה במספר הגידולים האינטסטינליים במודל השמנת יתר זה. התוצאות שלנו מעלות אפשרות נוספת, לפיה תכנית PPAR-δ בהנעת-HFD מזרזת בנוסף את הסוספטיביליות של non-ISCs לעבור טרנספורמציה אונקוגנית, ובכך לבסס מאגר תאים גדול ומגוון יותר המסוגל לאתחל גידולים. בהתאם לתפיסה זו, הוצע כי תאים ממוינים (non-ISCs) ברקע של מחסור ב- Apc עם אקטיבציה בו-זמנית של KRAS אונקוגני ואיתות NF-κB פרו-דלקתי הם בעלי יכולת אתחול גידולים. טרם ידוע האם דלקות הקשורות בהשמנת יתר ב- HFD תורמות לאיתות PPAR-δ וטומוריגנזיס אינטסטינלי, ובאיזה אופן. במודלים שלנו, לא מצאנו ראיות לכך ש- HFD או רכיבי חומצת השומן העיקריים שלה מביאים לאקטיבציה של נתיבים דלקתיים בקריפטים או אורגנואידים אינטסטינליים, בהתאמה (איור 3a-e extended data).
בעוד שמספר מחקרים קודמים מראים כי אינהיביציה של PPAR-δ עשויה לגרום להשפעות אנטי-סרטניות מתונות, אקטיבציה ממושכת של PPAR-δ ו- HFD נמצאו קשורות לאתחול והתקדמות של סרטן קולו-רקטלי. מחקרים עתידיים יצטרכו לבדוק האם אינהיביציה של PPAR-δ במסגרת של HFD משפיעה על האתחול וההתקדמות של גידולים. לבסוף, חשוב לחקור האם דיאטות קטוגניות רזות, שבדומה ל- HFD מורכבות בעיקר מחומצות שומן אך עם כמות קטנה יותר של פחמימות, מחקות את ההשפעות הפרו-רגנרטיביות של HFD תוך צמצום ההשפעות הבלתי-רצויות של השמנת יתר.
